Gastrulation Process in Frog and Chick Embryos

Gastrulation is a crucial stage in embryo development where the blastula transforms into a gastrula, involving the formation of the archenteron and primitive germ layers. Morphogenetic movements play a key role in this process, leading to the reorganization of cells. In frogs, gastrulation involves processes like invagination, blastopore formation, involution, epipoly, and rotation. Different types of morphogenetic movements, such as epipoly and emboly, contribute to the shaping of the embryo during gastrulation.

Download Presentation

Please find below an Image/Link to download the presentation.
The content on the website is provided AS IS for your information and personal use only. It may not be sold, licensed, or shared on other websites without obtaining consent from the author. Download presentation by click this link. If you encounter any issues during the download, it is possible that the publisher has removed the file from their server.
E N D
Presentation Transcript
MORPHOGETIC MOVEMENT& GASTRULATION IN FROG AND CHICK B.Bindu Bennet Assistant Professor, PG Department of Zoology Sarah Tucker College Tirunelveli
Introduction: The events which transform a blastula into a gastrula are collectively called gastrulation. According to McEven gastrulation refers to the formation of the promordial gastric or gut cavity called the archenteron. In addition to the formation of gastrular cavity, the process also involves the development of the two of the three primordial germ layers referred to as the ectoderm and endoderm. The third layer called mesoderm is derived from one or the other or both of the two layers, the endoderm is the inner layer, and the mesoderm is the middle layer.
Morphogenetic Movements : During gastrulation, the blastomers move from one place to another. These cell movements transform the blastula into another stage called gastrula. As these movements lead to the formation of a new shape to the embryo, these movements are called morphogenetic movements. Since the movements of cells from one place to the other bring about a new shape to the embryo, the movement are said to be morphogenetic movements.
Types of morphogenetic movements: The movements of the cells from one place to another are a common phenomenon in the embryos. These movements are of two types. They are Epipoly and Emboly. Epipoly : This word is derived from Greek, meaning throwing on or extending upon . It refers to the movements of cells on the surface of the embryo. Emboly : This word is also derived from Greek, meaning throw in or thrust in . It refers to the movements of cells from the surface to the interior of the embryo.
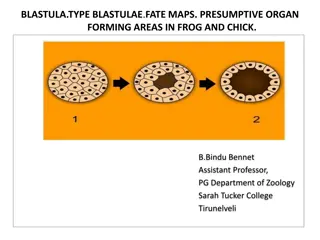
Gastrulation in Frog : The end of cleavage and the formation of the blastula is followed by the next phase of development called gastrulation. This is a phase during which the materials comprising the blastula are redistributed in such a way as to create a stratified embryo termed the gastrula. The events by which the blastula converted to a gastrula comprise gastrulation. Several processes occur simultaneously during gastrulation. It includes four kinds of morphogenetic movements, Invagination and formation of blastopore Involuation Ebipoly Rotation
Invagination and formation of blastopore : Gastrulation in frog is inaugurated by the formation of a depression or groove on the dorsal side of the embryo, at the boundary between the grey crescent and the ventral region. This groove represent the blastopore. It is prodeced by the invagination of the endodermal and mesodermal material into the interior of the embryo. These cells assume the elongated shape of a bottle and stream into the interior of the embryo toward the blastocoel cavity. Their steadily elongating necks remain attached to the surface of the blastula, whereas the bulky cell bodies move toward. As a result, a pull is exerted along their thin necks which creates an indentation at the surface. With continued multiplication and thinning out of the bottle cells, the indentation or invagination deepens. The area of the blastoderm immediately spoken of as the dorsal lip of the blastopore.
On the dorsal side of the embryo, a cavity is formed which leads from the groove on the surface into the interior of the embryo. Lined on all sides by the invaginated cells, this cavity is called archenterron. It is at the first narrow slit like cavity, but later, as more cells invaginate, it expends as its anterior end. In the process, it pushes the yolk-filled megameres toward the blastocoal and the latter is eventually obliterated. Involution and closure of the blastopore : In frog, invagination is always accompanied with involution. Involution can be described as the rolling of the superficial cells over the rim of the blastopore and into its interior while new portions of the blastopore approach the rim in their place.
Thus cells are carried inside of the gastrula, and tucked in beneath the outer layer. Involution is most active on the dorsal lip of the blastopore. The Prechordal plate: In the blastula it lies just below the presumptive notochord. The prechordal plate is the first mesoderm which involutes over the dorsal lip of the blastopore and becomes a part of the archenteron roof. In the gastrula, it lies in front of an anterior end of the presumptive notochord. The notochord : The notochord involutes by rolling over the dorsal lip of the blastopore into the interior of the embryo. It becomes stretched along the dorsal side of the archenteron and forms the mid-dorsal trip of the archenteron roof. During the process, the prospective notochord elongates in the longitudinal directions. However, it also undergoes a corresponding contraction in the transverse direction.
The mesoderm : Most of the mesoderm turns the corner and mocves to the interior by rolling over the lateral and ventral lips of the blastopore. After entering the embryo, the mesoderm advances in an anterior direction in the form of a sheet of cells and that penetrate between the ectoderm on the outside and the endoderm on the inside. Epiboly : Epiboly is the overgrowth of the ectoderm-forming regions around the inwardly moving prospective mesodermal and endodermal regions. The presumptive epidermis and the presumptive nervous system that are present in the animal region, greatly increase in their surface during gastrulation. This increase of surface area takes place at the expense of thinning out of the epithelial layer. The prospective epidermis expands in all directions, but in the case of the perspective nervous system the expansion is mainly in the longitudinal direction towards the blastopore.
As a result of epiboly, therefore, the ectoderm covers the whole embryo after the mesoderm and the endoderm have disappeared inside at the end of gastrulation. Rotation : Movement of the heavy yolky megameres from the posterior to the ventral position, reduction of the anterior blastocoel , and the enlargement of the dorsal archenteron together change the gravitational axis of the gastrula, shifting it about 90 away from the position of the blastopore. It causes the vegetal region of the embryo to be drawn into the interior and at the same time to rotate. Therefore, at the end of gastrulation, the vegetal region comes to lie in the ventral part of the archenteron, the embryo swings round within the vitelline membrane to bring its anteroposterior axis in a horizontal plane. Further, the presence of the archenteric roof and the pre-chordal and notochordal cells above it cause the overlaying ectoderm to thicken and the whole embryo becomes elongated in an antero-posterior direction.
The embryo is now called gastrula. The completed gastrula is nearly 3 layered . Endoderm is lacking only in the mid-dorsal region, but soon the free edges of the endoderm approach each other in the mid-line and fuse, thus completely enclosing the midgut. In this way, the three primary germ layers ectoderm mesoderm and endoderm are established. Gastrulation in chick : The process of gastrulation in the chick was studied by Chen, Hunt, Rawles, Rudnick, Woodsidde, Pasteels, Peter and Spratt. It is separable into three distinct phases: Laying down of the embryonic endoderm Formation of the primitive streak and mesoderm Formation of the embryonic axial and paraxial structures.
Formation of the endoderm : The first step in gastrulation in marked by the appearance within with the subgerminal cavity of a second layer, the hypoblast which, with later addition from the primitive streak, becomes the embryonic endoderm. After the formation of the hypoblast the upper layer is called the Epiblast which encompasses prospective ectoderm and the mesoderm. The space between the hypoblast and the underlying yolk then becomes the archenteron. There are different theories regarding the origin of hypoblast. They are as follows : Theory of involution : This theory was proposed by Patterson. According to this theory the hypoblast originates by involution of marginal cells through a temporary incontinuity in the zone of junction along a small part of the blastoporal ring.At present, this theory is not accepted.
Theory of invagination : According to this theory, proposed by Jacobson, the posterior end of the blastoderm invaginates into the subgerminal cavity to form a small pocket. The cavity of the pocket is the Archenteron and the floor and roof become the endoderm. This theory is also not entertained now-a-days. Theory of infiltration : This theory was proposed by Peter. According to this theory the individual cells detach themselves from the surface near the posterior end of the blastoderm. Peter was of the opinion that the process of infiltration took place mainly toward the future posterior side, with the subsequent forward movements. According Pasteels, it takes place more or less all over the area pellucid. Later, this sheet of embryonic endoderm becomes continuous with the extra embryonic endoderm around the margins of the blatoderm.
Formation of the primitive streak and mesoderm : After the separation of the endoderm, the next phase in gastrulation is concerned with the inward migration of the mesoderm and part of the endoderm in a special region inside the area pellucid. This area is the narrow strip of blastoderm which begins to appear at the posterior edge of the area pellucid. It gradually extends forward along the midline of the embryo and occupies about 3/5 th of the entire length of the area pellucid. This strip of gastrulation blastoderm is brought about by a convergence of the upper layer cells towards the midline. blastoderm becomes are known as the primitive streak. The thickening of the thickened during the stage of the The accumulation of cells is seen on the surface of the blastoderm as a dark or apaque streak, the primitive streak. It first becomes visible in the hindmost part of the area pellucid and is called the short primitive streak.
The mature primitive streak becomes narrower and quite sharply delimited. This is called the definite primitive streak. Formation of embryonic axial and paraxial structures: Head process or notochored process : The association of prospective notochord with the anterior end of the primitive streak is manifested by a noticeably thickening, the primitive knot or Henson s node. Henson s node is indicative of a special type of movement :the middle movement of cells. The prospective notochordal cells start moving as a dense mass in the middle straight forward from Henson s node underneath the surface of the epiblast. The tongue of notochordal cells is called the head process or the notochordal process. It can be easily distinguished from the surrounding cells. The established of the head process marks the completion of the elongation of the primitive streak.
The head process actually consists of a thicker central mass of cells and more diffuse lateral wings. The thicker central portion along gives rise to the definite notochord, whereas the lateral wings contribute to the paraxial mesoderm that comes to flank the notochord. The somites : The area of prospective somites lies in the anterior part of the primitive streak. From this part of the streak, the somite materials invaginates into the interior and becomes distributed in a strip on each side of the notochordal process. After formation of the notochord, the dorsal mesoderm becomes subdivided into segment the somites. The sementation proceeds from the anterior to the posterior end. It has been observed that one pair of somites is formed for every hour after 20 hours of incubation and the process continues unto about 40 hours regularly.
Therefore, an embryo with 4 pairs of somites is 24 hours old; an embryo with 16 pairs of somites is 36 hours old and so on. Thus, the total number of somites present at a given stage of the embryo is added to 20 to determine the age of chick embryo. The fully formed Chick gastrula has three germ layers : Ectoderm Chorda-mesoderm Endoderm.